Табл. 32 резюмирует другие результаты сравнения пород, выращиваемых в южных районах. Многочисленные сравнения кроликов средних пород Европы и Соединенных Штатов, таких как Новозеландская Белая и Калифорнийская, для которых Rochambeau (1988) составил резюме, были преднамеренно опущены. Табл. подчеркивает важность исследований, проведенных в Индии и Египте. К сожалению, мало исследований было проведено с местными популяциями. Новозеландские Белые и Калифорнийские кролики используются многими авторами, но это очень разные линии. Поскольку большинство авторов редко определяет происхождение животных для своих исследований, трудно сравнить результаты опытов между собой. Эти различные популяции белых кроликов могут иметь лишь один общий фенотип окраски. Эта табл. также показывает важность определенных популяций "гигантов", найденных в Индии и Египте, но, не зная размер взрослых животных этих популяций, невозможно определить, действительно ли они принадлежат к гигантским породам. Другие популяции, такие как Русская Шиншилла или Песчаный кролик, также заслуживают внимания.
Биологические компоненты плодовитости. Описание биологических особенностей местных популяций и пород дает полезную информацию для дальнейшего определения лучших стратегий использования. Для оценки плодовитости необходимо подсчитать количество желтых тел, чтобы оценить степень овуляции. Затем, чтобы определить эмбриональную жизнеспособность подсчитывается количество мест имплантации и количество живых и мертвых эмбрионов. Размер помета при окроле заканчивает оценку эмбриональной жизнеспособности. Наблюдая за рогами матки после имплантации эмбрионов (с 15-го дня беременности и до семи дней после окрола), можно оценить как степень овуляции, так и жизнеспособность эмбрионов. Самый простой метод – лапаротомия (вскрытие брюшной полости) с последующим исследованием яичников и матки. Поскольку обычно это требует умерщвления самки, то сегодня лучшим методом считается лапароскопия (диагностический осмотр брюшной полости и внутренних органов). Негативное влияние на самку значительно уменьшается при использовании эндоскопа, применение которого позволяет сохранять жизнь крольчихе и не нарушать ее воспроизводительную функцию. Табл. 33 и 34 показывают, некоторые различия между линиями. Линии отличаются изменениями между овуляцией и рождением, то есть линия 2066 отличается слабой предимплантационной жизнеспособностью (Табл. 33)
Табл. 33
 Табл. 34
Табл. 34
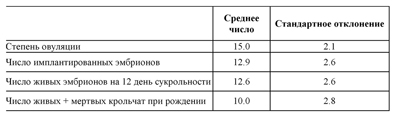
Источник: Santagreu, 1992.
Увеличение веса и скорости роста главных тканей зависят от биологических характеристик породы и от производственных факторов, таких как тип кормления. Так что критерием описания породы в определенных производственных условиях должна быть зрелость тела на единицу веса, определяемая как вес в данном возрасте, разделенный на взрослый вес.
С точки зрения производства мяса наиболее интересны породы с лучшим отношением прироста живой массы к взрослому весу, что позволяет таким кроликам рано доходить до надлежащего для продажи живого веса. Легковесные породы могут использоваться в виде чистых пород или в сочетании с породами среднего веса для получения молочных тушек с хорошо развитой мускулатурой и качественным мясом (достаточное количество жира) в соответствии с запросами потребителя.
Генетическое влияние на характеристики разведения
Генетическое улучшение характеристик разведения относится к определенной генетической изменчивости, выраженной в условиях той или иной окружающей среды. Эта изменчивость выражена как у животных одной породы или местной популяции, так и у различных пород и популяций и в межпопуляционных кроссах. Изменчивость - проявление генетических различий, которые отбираются и закрепляются путем скрещиваний с целью последующего использования.
Вопрос здесь стоит следующим образом: как генетическая изменчивость может использоваться на маленьких фермах с преимущественным использованием местных ресурсов? Повышение потенциала породы зависит от ее биологических характеристик, способности к воспроизводству и вычисления генетических параметров для селекции.
Биологические характеристики
Управляемое разведение. Один цикл размножения, требует много заботы и времени для получения первого и последующих окролов от самки. Крольчиха, обитающая в маточной клетке должна обслуживаться самцом в его клетке, на его территории. Теоретически, самка, достигшая сексуальной зрелости, может быть оплодотворена самцом в любое время, кроме периода беременности, но практически она не всегда допускает самца. Допуск самца с последующим спариванием дает потомство только в 70% случаев. Этот показатель изменяется в соответствии с физиологическими условиями, временем года, породой и окружающей средой. Рис. 17 резюмирует соответствующие генетические роли самца и самки, влияющие на размер помета при отъеме.
ТАБЛИЦА 35 Изменение живого веса молодняка кроликов в возрасте от 28 до 78 дней, и тушек, для двух пород
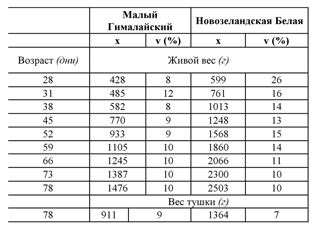
Внимание: Животные выращены в INRA (Тулузский центр). Применялась система рационального производства; отъем в 28 дней. Тушки с головой и лапками. x = среднее число; v = коэффициент изменчивости.
ТАБЛИЦА 36 Средний живой вес в 84 дня, вес тушки, отношение мышцы/кости, вес жировой ткани в тушке для трех пород
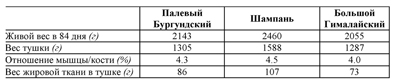
Источник: Rouvier, 1970.
Первый значительный шаг к проведению спаривания зависит от сексуального возбуждения самца и самки. Немного известно о биологических основах сексуальности кролика. Сексуальная функция снижается при высокой температуре окружающего воздуха (28 - 30° C). В жаркие времена года самка должна быть представлена самцу рано утром, в 6.00 или еще раньше, когда сексуальное возбуждение самое большое.
На плодовитость влияет, во-первых - овуляция, которая зависит от самки и происходит через 10 часов после спаривания и, во-вторых - оплодотворение яйцеклетки, что зависит от самца и происходит через 16 часов после спаривания. Гены, как самца, так и самки одинаково влияют на внутриутробный рост и жизнеспособность зародыша. Кроссирование может улучшать жизнеспособность яйцеклетки, бластоцисты и эмбриона. Самка имеет большее влияние на рост и развитие зародышей в утробе, поскольку это обусловлено в большей степени питанием эмбриона. Самец же может оказать влияние на количество крольчат в окроле.
РИСУНОК 17 Соответствие генетических ролей самца и самки кроликов в определении размера помета при отъеме
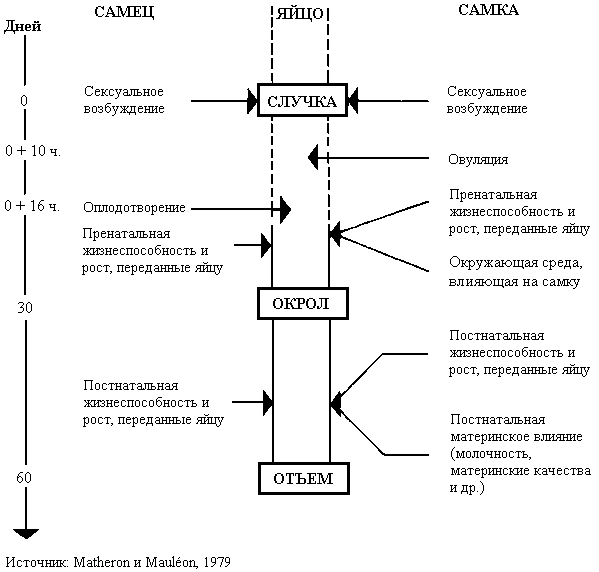
Плодовитость самки – породная характеристика, но с существенными индивидуальными колебаниями (от 1 до 18 крольчат в окроле). После окрола самки весь помет должен быть благополучно выращен до срока отъема. Селекционер влияет на размер помета до отъема, защищая молодняк и обеспечивая кормом кормящую самку. Жизнеспособность крольчат, материнское поведение и секреция молока также важны. Жизнеспособность в подсосный период зависит от количества живых крольчат при рождении, как показано в табл. 37.
Показатель жизнеспособности остается довольно постоянным при количестве новорожденных в окроле от трех до девяти. Маленький окрол (один или два живых новорожденных) не может создать благоприятную окружающую среду для выживания крольчат. Резкое снижение жизнеспособности молодняка в подсосный период наблюдается при показателе 8.57 крольчат, от окролов по 12 и более крольчат. Это подталкивает на осуществление работ, связанных с пересадкой крольчат из одного гнезда в другое гнездо. Целью проведения подобных мероприятий является увеличение общего выхода молодняка ко времени отъема. Кролики-подкидыши могут прибывать из малоплодных гнезд (один или два крольчонка), или, что более обычно, из многоплодных окролов (более десяти крольчат). Однако пересадка крольчат из одного гнезда в другое подразумевает достаточное количество самок в крольчатнике и наличие у них очень хороших материнских качеств. В день окрола, сразу же после первого кормления крольчонок может быть отделен от матери на период до 24 часов, что может предполагать небольшое путешествие и перемещение в гнездо к другой крольчихе-кормилице.
ТАБЛИЦА 37 Жизнеспособность крольчат до отъема в зависимости от размера окрола
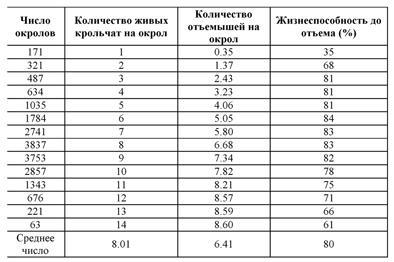
Обратите внимание: Данные получены от рациональной фермы, расположенной Юге Пиренеи во Франции.
Источник: Roustan, Matheron и Duzert, 1980.
Биологические характеристики крольчихи, такие как овуляция, вызываемая спариванием, принятие самца на следующий день после окрола, отсутствие лактации в анэструсе (период отсутствия течки), отсутствие сезонного анэструса позволяют рассматривать широкий диапазон теоретических темпов воспроизводства. Как пример, табл. 38 сравнивает три различных варианта темпов воспроизводства в коммерческом центре разведения кроликов в Мексике.
Как самки, так и самцы имеют очень высокий потенциал воспроизводства, что подтверждается третьим опытом из табл. 38. Потенциальное воспроизводство на самку в год может оцениваться в 150 крольчат. Однако для достижения этого уровня производства потребуется много лет исследований, а также мастерство управления факторами окружающей среды. В настоящее время в развивающихся странах при разведении кроликов стремятся использовать местные популяции и более длинные репродуктивные циклы. Улучшение технического оборудования должно начаться на базе модернизации традиционных методов производства и (где они существуют) местных популяций.
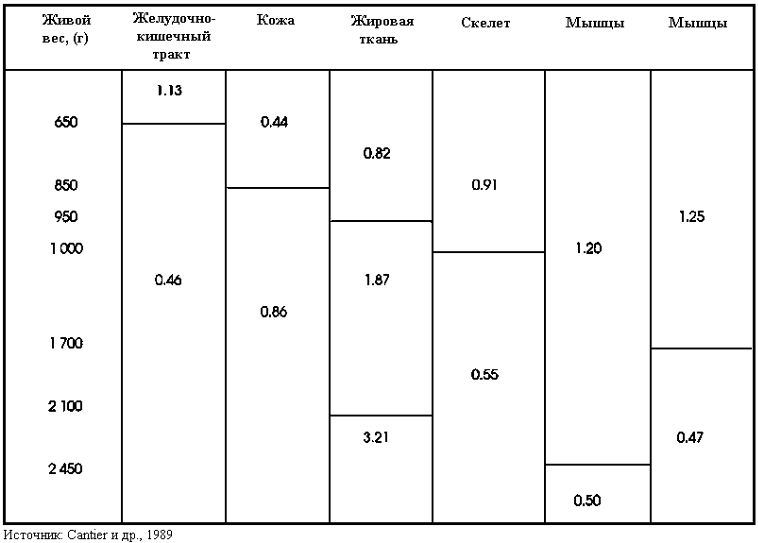
Рост тканей. По исследованиям Cantier и др. (1969), у кроликов в первую очередь развивается костная ткань, затем к развитию скелета присоединяются мышечная ткань, затем жировая ткань. У популяции обычных кроликов со средним взрослым весом (4 кг) скелет развивает быстро до достижения живого веса 900 г. Затем рост скелета замедляется вплоть до достижения кроликом взрослого веса. Мышечная ткань очень быстро наращивается до достижения живого веса 2.3 - 2.6 кг, затем кривая роста резко падает. Жировая ткань развивается быстрыми темпами после достижения живого веса 2.1 кг. Учитывая различную скорость роста отдельных тканей по отношению к взрослому весу необходимо регулировать рацион в зависимости от половозрастной группы и породы. В итоге, для извлечения максимальной выгоды кролики должны отправляться на убой имея 50 - 60% от нормального взрослого веса, характерного для той или иной породы или популяции. Это наиболее оптимальная степень развития кролика для получения тушки с лучшим анатомическим составом и наиболее эффективным использованием корма.
ТАБЛИЦА 38 Сравнение трех норм воспроизводства

Источник: Национальный Центр Кролиководства Ирапуато, Мексика.
Неполноценный корм замедляет рост и развитие и снижает конверсионную эффективность - количество корма, необходимого для производства 1 кг привеса. Это не является проблемой, если система разведения предусматривает кормление молодняка кроликов с помощью местных ресурсов, но как показывает практика, из всей популяции, наиболее быстро растущие животные к возрасту убоя имеют лучший вес и состав тушки (отношение мышцы/кости, процент жира). Естественно, мясо молодого кролика более постное; нет никаких излишков жира. Лучший возраст и вес для убоя должен быть разработан на основе рыночного спроса, системы производства и типа кормления.
Гены и окружающая среда
Наиболее количественные характеристики размножения - плодовитость, жизнеспособность, рост, и т.д. устанавливаются генетически, но они также подвержены влиянию окружающей среды. Фенотип - результат воздействия генотипа и окружающей среды в виде определенных характеристик. Генотип - результат воздействий генов в нескольких локусах. Окружающая среда состоит из множества компонентов: климат региона, среда обитания, микроклимат, окружающий животных, температура, влажность, скорость воздуха, оборудование крольчатника, методы разведения, методы кормления, и человеческий фактор - кроликовод. Определение разновидностей генетических характеристик имеет двойной интерес для селекционера и кроликовода: во-первых, чтобы использовать генетическую изменчивость животных той же самой породы или популяции; и, во-вторых, методом скрещиваний эксплуатировать генетическую изменчивость между породами и популяциями.
Индивидуальная генотипическая ценность непосредственно не заметна, и только ее проявление (фенотип) может быть измерено. Обычная модель количественной генетики предполагает, что фенотипическая ценность будет складываться из суммы генетической ценности и факторов окружающей среды. Эта модель рассматривает генотип и окружающую среду, как два независимых компонента. Согласно этой модели, генетическая ценность - частично результат совокупной генетической ценности и частично воздействие гена на тот же самый локус (доминирование) или другие локусы (эпистаз). Используя коэффициент регрессии, совокупная генетическая ценность индивидуума оценивается по результатам продуктивности этого индивидуума и его родственников. Целью селекционной программы является создание генетического прогресса, то есть увеличение средней совокупной генетической ценности популяции.
ТАБЛИЦА 39 Аллометрические коэффициенты главных органов и тканей и указание критического живого веса (меньше содержимого пищеварительного тракта) наблюдаемые у самцов
Наследственность и генетические корреляции. Уровень генетического прогресса зависит, прежде всего, от того какова величина колебания совокупного генетического происхождения. Этот коэффициент был назван наследственностью, и рассчитывается он как отношение совокупного генетического колебания к полному колебанию. Поэтому наследственность изменяется от ноля до одного. Также, наследственность - коэффициент регрессии совокупной генетической ценности индивидуума по его/ее собственным результатам работы. Наследственность изменяется со степенью изучения популяции и окружающей среды. Особенно она варьирует в отношении частоты гена и таким образом изменяет ее в селекционируемой популяции.
Рис. 18 показывает наследственность основных характеристик интересующих селекционеров. Наследственность читается по часовой стрелке слева. Фертильность самки около нуля в отношении наследственности. Затем, перемещаясь по часовой стрелке, наследственность увеличивается. Наследственность размера окрола - приблизительно 0.10. Самый высокий коэффициент наследственности у живого веса в возрасте отъема и в 77 дней (0.2 - 0.3) и этот показатель увеличивается, поскольку животное растет и материнское влияние уменьшается. Скорость роста в послеотъемный период и эффективность кормления в групповых клетках колеблются между 0.3 и 0.4. Коэффициент более 0.4 имеют такие характеристики как вес тушки, соотношение мышцы/кости, потребление в групповых клетках и убойный выход.
РИСУНОК 18 Наследственная и генетическая корреляция производственных характеристик у кроликов
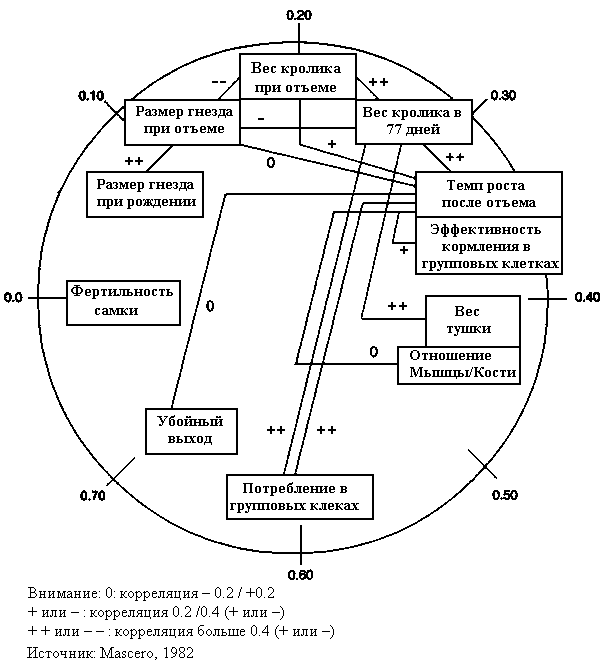
Эти оценки относительны, поскольку проблема оценки вариантного отношения на доступных данных заключается в том, что исходные данные составляются с учетом наследственности в данном месте и данное время. Обзор Rochambeau (1988) - поучительная иллюстрация этого: наследственность для количества живых новорожденных крольчат изменяется от 0.0 до 0.4, когда верхняя и нижняя четверть оценочных показателей удалена. Разброс для индивидуального веса в 14 недель - 0.2 и 0.8 при тех же самых условиях.
Генетическая изменчивость среди пород и популяций. Местные породы или популяции могут сравниваться с улучшенными породами других странах или с кроликами, выращенными в различных производственных системах. Различия в породах, прежде всего, закрепляются через межпородное скрещивание животных (кроссбридинг). Поэтому сравнительная характеристика между породами в крольчатниках очень полезна. Местные породы и популяции могут сравниваться с улучшенными породами других стран и породами, произведенными в различных условиях. Межпородные различия эксплуатируются в основном через кроссы. Не все кроссы выгодны, однако; все они должны быть проверены. Главные преимущества кроссбридинга - гетерозис и удачная сочетаемость пар при скрещивании.
Понятие гетерозиса может быть определено как получение лучших показателей продуктивности при кроссе, чем эти же усредненные показатели двух чистых родительских пород. Гетерозис может отражаться на молодняке (например, повышается его жизнеспособность), на самке (плодовитость, молочность) или самце (энергичность, сексуальное возбуждение, плодовитость). Доминантные характеристики, такие как характеристики воспроизводства, являются наиболее вероятными, для извлечения выгоды из гетерозиса.
При скрещивании популяций отличающихся генетически гетерозис может иметь место, но он не всегда будет проявляется путем изучения фенотипически чистых пород или популяций. Помесные животные всегда в большей степени разнородны, чем животные, полученные от двух родительских популяций, и это подразумевает большую адаптируемость к изменениям и суровым условиям окружающей среды. Поэтому, кроссирование может быть полезно в случае, если целью является получение кролика с улучшенными характеристиками для последующего использования в развивающихся странах, но все скрещивания должны быть запланированы. Там, где существуют местные популяции кроликов, использование кроссбридинга рекомендуется.
Кроссбридинг делает возможным оптимальное использование удачного сочетания пар при скрещивании пород или популяций. Удачное сочетание пар при скрещивании касается двух групп - показатели матери и ее молодняка, которые влияют на суммарный выход мяса, произведенный самкой. При кроссбридинге удачное сочетание пар направлено на соединение производственных характеристик матери и потомства, или на благоприятную комбинацию совокупных эффектов этих характеристик.
В первом случае, самцы одной породы с высоким потенциалом роста скрещиваются с самками другой породы или популяции, которые показывают хорошие показатели плодовитости, высокие материнские качества и неприхотливость к условиям окружающей среды. Второй случай касается характерных особенностей, составляющих полную характеристику. Таким образом, количество овуляций, оплодотворившихся яйцеклеток и жизнеспособность эмбрионов - компоненты, которые в итоге влияют на размер помета при окроле (плодовитость). Плодовитость и жизнеспособность до отъемного возраста - компоненты, влияющие на размер помета при отъеме. Поэтому должны быть найдены такие скрещивания, которые объединили бы высокий уровень овуляции и хорошую жизнеспособность эмбрионов у скрещенной самки. Эти характеристики могут быть хорошими антагонистами на внутрипопуляционном основании.
Эффект гетерозиса и удачное сочетание пар носят не систематический характер. Программы кроссирования необходимы для ясного обнаружения этих эффектов. Рассмотрим это на примере популяции А и популяции B. Рекомендуется, чтобы обе популяции были чистокровными (A × A) и (B × B). Это необходимо для сравнения их с двумя кроссами (A × B) и (B × A), для выдвижения на первый план показателей материнских и праматеринских поколений.
Для интуитивной иллюстрации материнского эффекта, предположим что, порода A имеет взрослый вес 6 кг, а породы B взрослый вес 3 кг. Мы скрещиваем самца A с самкой B и самца B с самкой A, и сравниваем вес молодняка при отъеме. Молодняк AB имеет тот же самый генетический набор, что и молодняк BA, поскольку они образованы на половину из отцовских и на половину из материнских генов. Однако молодняк получает разный материнский уход: самки A имеют большую матку и производят большее количество молока, так что их молодняк имеет больший вес при отъеме. Таким образом, имея том же самый генетический набор, BA кролики более к сроку отъема имеют больший вес, чем AB кролики из-за влияния более благоприятного материнского ухода. Более точное определение дали Matheron и Mauléon (1979). Они рекомендуют, чтобы как минимум два поколения кроссов были изучены, с целью обнаружения прямого эффекта гетерозиса, проявляющегося на производственных показателях кроликов и на материнских эффектах. Первое поколение включает скрещивания (A × A), (B × B), (A × B) и (B × A); второе поколение включает скрещивания чистых самок AA и BB и помесных самок AB и BA с самцами линии C. Если изучено более двух популяции, количество генотипов для сравнения во втором поколении уменьшается с квадратом количества популяций.
В качестве примера рассмотрим эксперимент, проведенный в Тулузском Центре INRA в 1987-1989. Этот эксперимент, проводимый в три стадии, использовал линии 1077, 9077 и 2066, как показано в табл. 28. Первая стадия включает спаривание между самцами и самками трех линий: самцы каждого генотипа (1077, 9077 и 2066) скрещивались с самками каждого генотипа (1077, 9077 и 2066) для получения девяти пометов с разными генотипами (три чистых и шесть кроссированных). Во второй стадии, самки этих девяти генотипов случались с самцами трех чистых генотипов. В последней стадии, те же самые самки были случены с самцами, принадлежащими к двум линиям последнего кроссирования различного происхождения. Первые три помета были исследованы в каждой стадии. Самки были убиты во время их четвертой беременности, и затем составляющие размера помета были изучены.
Табл. 40 сравнивает показатели, полученные от чистых и кроссированных самок. В целом, показатели кроссированных самок были выше, количество овуляций было выше на 1%, а к отъему размер гнезда был выше на 13%. Также существуют различия между результатами, полученными от чистых линий и кроссированных самок. Последующий анализ полученных данных объясняет различия между скрещиваниями для дальнейшего использования. Самки 2066 имеют лучшую степень овуляции, но это преимущество исчезает в следующей стадии. Показатели продуктивности 2066 и 1077 довольно схожи. Результаты работы линии 9077 менее хороши.
Генотипы, кроссированные с линией 2066 также имеет более высокую норму овуляции: преимущество прослеживается вплоть до отъема, где генотипы 2066 × 1077 и 1077 × 2066 подтверждают свое превосходство. Размер помета значительно увеличился за счет кроссированных самок.
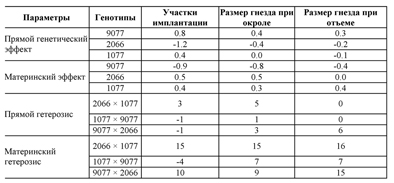
Табл. 41 анализирует те же самые результаты в показателях генетического эффекта. Оценка прямого генетического эффекта показала, что линия 2066 имеет отрицательный эффект по количеству участков имплантации, а линия 9077 имеет положительное воздействие на размер помета при окроле. Материнское влияние линии 9077 по количеству участков имплантации контрастирует с положительным эффектом линии 1077 по размеру помета при отъеме. В то время как эффект прямого гетерозиса слабый, материнский гетерозис, оказывает главный эффект на количество участков имплантации и поддерживается вплоть до отъема и достигает 16% для линий 1077 и 2066.
Результаты экспериментов по кроссированию имеют определенный интерес для выбора оптимальной стратегии селекции. Особенности популяции животных изучаются и не могут быть обобщены для всех животных в породе. Однако полученные данные могут описывать местные популяции или линии, таким образом, выбирая лучший способ использования их в кроссбридинге или чистом разведении.
Кроссбридинг в тропических странах. Биологическая основа для будущих кроссов должны быть найдены среди доступных популяций животных, разводимых в условиях различных окружающих сред. Несколько крупномасштабных исследований межпородных кроссов в тропических странах были проведены на Кубе, и в Египте.
Эксперимент проводился на импортированных кроликах акклиматизированных пород, а не на местных популяциях кролика. Используя лучшие кроссы, была улучшена мясная продуктивность. В 1969 - 1971 г.г. Кубинский Институт Научного Животноводства, основываясь на чередовании, провел работу по скрещиванию четырех пород: Белый Полугигант, Калифорнийская, Новозеландская Белая и Шиншилла. Анализировались следующие характеристики: размер гнезда при окроле и отъеме, и вес гнезда при отъеме. Эксперимент проводился в течение сухого сезона (с ноября до апреля, средняя температура 22.2 °C, влажность 75.2%) и сезона дождей (температура 26.1 °C, влажность 77.7%). Некоторые из экспериментальных животных этих четырех пород недавно были импортированы из Канады, другие уже находились на Кубе в течение некоторого времени. Взрослый живой вес приводится в табл. 42.
ТАБЛИЦА 40 Средние показатели продуктивности самок девяти генотипов: составляющие размера окрола, измеренные в различных стадиях
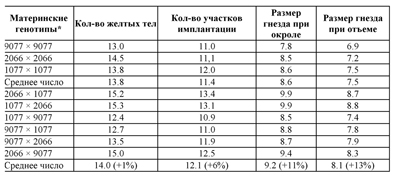
* Отцовский генотип, сопровождаемый материнским.
Источник: Brun, Bolet и Ouhayoun, 1992.
ТАБЛИЦА 41 Генетические параметры размера гнезда, измеренные на различных стадиях между овуляцией и отъемом
Источник: Brun, Bolet и Ouhayoun, 1992.
Животные выращивались в клетках, идентичных тем, что используются для выращивания кроликов в южной Калифорнии. Это сетчатые клетки с деревянными гнездовьями, расположенными в отдельных углублениях в два ряда, покрытых крышей зданиях, открытых со всех четырех сторон. Эта среда обитания защищает кроликов от прямых солнечных лучей, но во влажном тропическом климате не может защитить от дождя и ветра, что и объясняет высокую смертность кроликов до отъемного возраста.
Использовалась экстенсивная система воспроизводства, с отъемом в 45 дней, отъем молодняка сопровождался спариванием самки. Средняя плодовитость для пород этого взрослого размера (общее количество крольчат в помете - 7.45) в пределах нормы; среднее значение мертворожденных в окроле слегка превышает норматив (более чем 10%); и, прежде всего, высокая смертность в период от рождения до отъема (2.5 отъемыша на помет). Это было вызвано несоответствующей защитой гнезд от ветра и дождя, и неадекватным кормлением лактирующих самок. Положительный вклад кроссбридинга в таких трудных условиях производства представляет определенный интерес.
Сравнительный анализ результатов среди чистых пород показал, что Белый Полугигант теряет меньшее количество молодняка между окролом и отъемом, чем другие породы, и вес отъемышей лучше. Для простых кроссов самые высокие показатели по количеству отъемышей и самые низкие показатели смертности были зарегистрированы у кросса Новозеландская Белая × Белый Полугигант. Также, количественная продуктивность может быть увеличена путем скрещивания женского потомства, полученного в результате этого кросса с Калифорнийскими самцами. Наиболее продуктивный - самки Белый Полугигант × самцы Шиншиллы.
Afifi и Khalil (1992) обобщили результаты девяти Египетских экспериментов, изданных между 1971 и 1990. Они сравнили чистых и помесных животных от местных или импортированных популяций. Список используемых пород: Бускат, Шиншилла, Белый Гиза, Белый, Красный и Желтый Балади, Серый Фламандский Гигант, Белый Фламандский Гигант, Новозеландская Белая и Калифорнийская. Исследования включали много простых кроссов, но, к сожалению, мало помесных самок. Резюмируя, авторы делают вывод, что местные породы (Белый Гиза, Балади) превосходят по характеристикам, которые выражаются до окрола, а импортированные породы (Новозеландская Белая, Калифорнийская, Бускат) лучше показывают результаты послеродовых характеристик. Обзор включает много оценок эффектов прямого гетерозиса, показанных в табл. 43. В экспериментальных окружающих условиях, прямые эффекты гетерозиса оказались слабыми по параметрам изучения. Кроме средней ценности в 15% для веса молодняка при окроле и 7% для размера помета при отъеме, все другие ценности были ниже 5%. Они близки к нулю для индивидуального веса в 4 и 12 недель и для жизнеспособности в послеотъемный период. Эффект материнского гетерозиса более сильный, даже притом, что небольшое количество экспериментальных результатов не позволяет делать это заявление категоричным.
Улучшение генетических качеств: селекция и кроссирование
Юго-западные страны Европы - Франция, Италия и Испания развивают программы улучшения генетических качеств кролика для удовлетворения потребностей интенсивного производства в умеренном климате. Животные, селекционированные в Западной Европе, не обязательно являются лучшим вариантом для маленькой кроликофермы (5 - 60 самок) из-за различных производственных условий. Местные породы кроликов, разводимые в пределах определенной местности путем скрещивания с различными импортированными популяциями должны использоваться для улучшения генетических качеств.
Эффективное улучшение генетических качеств должно быть под руководством группы, имеющей научно-техническую поддержку в лице научно-исследовательских организаций страны. Программа улучшения генетических качеств должна сосредотачиваться на деревне (или предпочтительно на группе деревень), на всех крольчатниках региона, или на целой стране. Улучшение генетических качеств - дорогостоящая операция: рабочая группа должна быть достаточно большой, поэтому необходимо учесть покрытие связанных с этим расходов и мобилизовать необходимые профессиональные знания.
Улучшение генетических качеств требует специализированных технических знаний. Поэтому, должны быть кролиководы-селекционеры, кролиководы-пользователи и, возможно, кролиководы-репродукторы между ними. В то время как пирамидальные схемы, используемые в Западной Европе эффективны в их специальном контексте, они не универсальны в практическом применении. Каждая схема верна для индивидуальных условий региона, страны. Немаловажную роль в этих схемах играет социальная специфика кролиководов страны (хотя в любом случае, схемы должны быть генетически эффективны). Селекционеры также должны быть превосходными кролиководами, чтобы умело использовать производственные системы, кормовые ресурсы, размещение животных в помещении и другие материалы, взаимодействующие с окружающей средой. Сложных методов селекции не