
Введение
Домашние кролики – потомки диких кроликов из рода Oryctolagus, представленного видом cuniculus. Это аборигенный вид, обитающий на западе Средиземноморского бассейна (Испания и Северная Африка). Существуют также другие дикие кролики, принадлежащие к родам: Sylvilagus, Coprolagus, Nesolagus и Brachylagus. Кролик был одомашнен относительно недавно: большинство пород создано людьми и многим из них не более 200 или 300 лет, которые адаптированы к определенной местности.
Таксонометрическая классификация кролика (Oryctolagus cuniculus)
КЛАСС: MЛЕКОПИТАЮЩИЕ
- Надотряд: Glires
- Отряд: Lagomorpha (Зайцеобразные)
- Отряд: Rodentia (Грызуны)
Семейство: Leporidae (Зайцы)
+ Подсемейство: Palaeolaginae
- Род: Pentalagus. Восточная Азия
- Род: Pronolagus (Курчавохвостые кролики). Юго-восточная Африка
- Род: Romerolagus (Бесхвостые кролики). Мексика, один единственный вид, R. nelsoni
+ Подсемейство: Leporinae
- Род: Lepus. Зайцы: множество видов, распространенных по всему Старому и Новому Свету
- Род: Macrotolagus. Иногда рассматривается как подрод Lepus, живущий в Северной и Центральной Америке
- Род: Oryctolagus. Истинный дикий кролик, живущий в Европе и Северной Африке. Представлен одним видом - Oryctolagus cuniculus, с несколькими подвидами
- Род: Sylvilagus (Американские кролики). Представлен множеством видов
- Род: Caprolagus (Азиатские жесткошерстные кролики)
- Род: Nesolagus (Полосатые кролики). Суматра, один единственный вид, N. netscheri
- Род: Brachylagus (Карликовые, или Айдахские кролики). Это кролик-пигмей, живущий в Северной Америке
Семейство: Ochotonidae (Пищуховые)
- Род: Ochotona (Пищухи). Один единственный род для различных пищуховых; обитают в северных регионах Европы, Азии и Америки
Источник: Grassy и Dekeyser, 1995.
Кролик использовался как экспериментальное животное в генетике и физиологии воспроизводства начиная с начала столетия, но первые результаты по количественной генетике были изданы только в 1950. Это были исследования Венга, направленные на изучение материнского влияния на вес новорожденных кроликов (Венг, 1950). Эта работа проложила путь к исследованию улучшения генетических качеств кролика для производства мяса. В Национальном Институте Сельскохозяйственных Исследований (INRA) во Франции ученые начали проводить исследования в этой области в 1961. Далее к работам в этой области присоединились другие научно-исследовательские лаборатории во многих странах, таких как команда Университета г. Эз-Заказик в Египте, Годоло и Капошвар в Венгрии, Изванагара в Индии, Mилана и Витербо в Италии, Валенсии, Сарагосы и Барселоны в Испании. Хорошие команды в Соединенных Штатах и Китае (особенно в Шанхае) и определенная работа была проделана в Нитре в Словакии и в Кракове в Польше. Превосходная библиография Робинсона (1958) Об изучении генетики кролика, основанная на нормальных генетических и физиологических данных, теперь является устаревшей в свете новых исследований этих команд.
Работы по генетике кролика регулярно обновлялись на мировых конгрессах кролика (Rouvier, 1980; Matheron и Poujardieu, 1984; Rochambeau, 1988). Однако опыт, полученный в производственных условиях Европы, не может быть перенесен непосредственно в развивающиеся страны. Чтобы улучшить генетику кроликов таких стран, селекционеры должны использовать местных, или аборигенных животных, или кроликов ранее импортированных популяций, которые адаптировались к местным условиям, и используют доступную генетическую изменчивость.
Очевидно, что приоритет нужно отдать исследованию сельского и подворного производства кролика. Это были бы маленькие, экономные, автономные фермы, требующие небольших инвестиций и предусматривающие использование местных ресурсов. Такие фермы были бы разумно производительны.
Генетика пород кроликов и популяций
Наверное, лучшее из различных определений породы дал Квиттет: "Порода - группа индивидуумов в пределах вида, различающихся несколькими морфологическими и физиологическими признаками, которые они передают своему потомству, размножаясь между собой".
Единственный путь оценки генетической уникальности различных пород заключается в том, чтобы изучить их происхождение. Порода - результат совместного воздействия искусственного и естественного отбора (адаптация к окружающей среде). Трудно определить точно, что является породой, и какова ее предыстория. Искусственный отбор может быть направлен на ряд различных критериев, не обязательно все они связаны с производительностью. Условия разведения могут быть или искусственными или естественными, окружающая среда может постепенно изменяться и т.д.
Породы кроликов или популяции также могут быть описаны в терминах частоты гена. Идентификация и манипуляция возможны лишь с генами, которые проявляются на потомстве через видимые эффекты. Окраска и структура волос классифицируются как видимые эффекты. Благодаря современным методам изучения генов, теперь также известны гены, которые отвечают за разделение на группы крови, биохимический и белковый полиморфизм и наследственные аномалии. (См. Zaragoza и др., 1990.)
Количественные характеристики, такие как рождаемость или вес крольчат при отъеме, которые управляются очень многими неизвестными генами и могут быть учтены только после измерения выполненной ими работы. Эти гены имеют небольшой эффект на полную изменчивость и функционируют независимо, согласно стандартным положениям о количественной генетике. Такие характеристики находятся под влиянием окружающей среды. Характеристики окружающей среды должны быть тщательно описаны (количество кролиководов, направление селекции, происхождение популяции и ее распространение) при описании популяции.
Гены входят в структуру хромосом, которые объединены в 22 пары (2n=44). Описано приблизительно 60 маркеров. Изучаются гены с видимым эффектом, такие как окрас, длина или морфологические аномалии волосяного покрова, или кодирующие гены для молекул, которые исследуются на биологическое воздействие. Эти два подхода трудно урегулировать, поэтому часто используют только один тип маркера. Среди описанных маркеров, 37 были помещены в восемь аутосом и в хромосому X; 23 маркера составляют шесть групп сцепления, а месторасположение шести маркеров все еще не найдено. Все эти маркеры расположены в 22 парах хромосом кролика. Однако связи между биологическими маркерами и генами окраса или длины волос редко исследовались.
Опыты показали, что кролики могут выдерживать медленное и постепенное увеличение степени инбридинга, но в результате проведенных исследований ученые рекомендуют составлять план случек для маленьких популяций так, чтобы минимизировать степень его увеличения среди животных (Rochambeau, 1990).
Породы, созданные селекционерами, особенно любителями, в Соединенных Штатах и Европе, теперь соответствуют официальным стандартам. Книга Федерации французских кролиководов (FFC) по стандартам для кроликов описывает больше 40 пород. Каждую породу вывели из местных кроликов, или путём скрещивания существующих пород, или используя мутантов с измененной окраской или структурой меха. Массовая селекция по размерам и строению тела разделила эти породы на следующие типы: великаны, средние, маленькие и очень маленькие. Интересно изучение происхождения пород с целью обучения и выяснения причин того, могут ли они соответствовать первоначальным генетическим ансамблям и пытаться определять их характеристики.
Характеристики, которые характеризуют животное как соответствующее стандарту породы, такие как размер тела, действительно ли оно компактное, окрас волосяного покрова и плотность, размер ушей, могут быть связаны с сопротивлением организма изменяющимся условиям климата. Фактически, такие факторы как волосяной покров, кожа, площадь тела и вес влияют на температуру тела животных.
В настоящее время известны генетические детерминанты, отвечающие за окрас и структуру волосяного покрова. Более подробно они описаны ниже. Окраска волосяного покрова всегда имела большой интерес у селекционеров.
Окрас и структура волосяного покрова
В труде генетика домашних кроликов, изданного в 1930, Кастл описал шесть мутаций окраса волосяного покрова и две мутации манеры поведения; три мутации в структуре волос; одна мутация в желтой окраске брюшного жира и двух групп сцепления. Удобный способ обнаружения различных мутаций заключается в описании "дикой" окраски кролика. Волосяной покров состоит из трех типов волос: ретрикс - более длинный остевой волос, жесткий у основания; текрикс - более многочисленные покровные волосы, формирующие основную часть волосяного покрова; и третий типом - более короткие волосы, составляющие подшерсток.
Окрас дикого или "агути" кролика формируется из серого меха на спине и из намного более светлого или белого меха на брюхе. Длинные охранные волосы черные, но кажутся еще более черными на концах и более синими у основания. Колючие волосы имеют цветовую зональность: черные на концах, с желтой полосой в середине и синеватые у основания. Волоски подшерстка синеваты у основания и украшены бахромкой с желтыми кончиками. Таким образом, объясняется неодинаковая окраска тела кроликов из-за неравномерного распределения черных и желтых пигментов (меланины), особенно в колючих волосах, и в целом по всему волосяному покрову (бока и спина относительно меха на животе). Мутации в различных локусах (месторасположение гена в хромосоме) изменяют эту окраску.
Окраска. Обозначается в соответствие с международный системой обозначения аллелей. Арнольд (1984) привел обозначение окраса кроликов в соответствии с Немецкой системой.
- Локус A, агути: мутация a производит животных без желтой полосы в волосе и с более светлым животом. Их окраска однородна. Ген A является доминантным над a. Треть аллели была описана в этом локусе, at (рыжевато-коричневый образец), который является рецессивным к A и доминантным к a.
- Локус B, черный пигмент: рецессивная b аллель производит в волосяном покрове агути шоколадно-коричневый пигмент вместо черного пигмента.
- Локус C: ген C требуется для развития пигментов в мехе, коже и глазах и, следовательно, для выражения окраски. Рецессивный c ген подавляет выраженность окраски, порождает альбинизм в рецессивных cc гомозиготах. Существует несколько аллелей в этом локусе, указанных ниже в доминирующе-рецессивном порядке:
C: Полное выражение окраски.
сch : Шиншилла. Замена цвета промежуточной полосы волосяного покрова.
ch: Гималайский. Только кончики волос тела и конечности черного цвета. Степень проявления этого гена зависит от окружающей температуры.
c: Альбинизм. Локус альбинизма эпистатичен по цвету локуса. Генотип cc подавляет проявление генов цвета, расположенных в другом локусе.
- Локус D: рецессивная мутантная d аллель затрагивает интенсивность пигментации, вызывая растворение гранул пигмента. Доминирующая D аллель производит нормальную интенсивность пигментации. Рецессивная dd гомозигота найдена в генотипе синих (черные, растворенные до синего) или бежевых (желтые, растворенные до бежевого) кроликов.
- Локус E: мутации гена e служат причиной увеличения желтого пигмента в волосах, с тенденцией замены черного (или коричневого) пигмента. Серые, черные или коричневые породы имеют E ген. Желтые и красные породы – рецессивные ee гомозиготы.
- Локус V Венский Белый: Венские Белые кролики имеют полностью непигментированный мех, но цветные (синие) глаза. Первоначальный ген обозначается V, а его видоизмененная форма v. Поэтому кролики породы Венский Белый – рецессивные vv гомозиготы. Скрещивание этой породы с кроликами альбиносами дает цветное потомство.
- Мутации, дающие пятнистый окрас: эти мутации содержат локусы Английский ( En , en) и Голландский (Du, du) Кролик породы Бабочка имеет En en гетерозиготный генотип. Ген En не полностью доминирующий. En En гомозиготы более белые, чем гетерозиоты, в то время как рецессивные гомозиготы более черные. Окрас генотипа кролика Бабочка (Великан Чекер в Англии, Марипоса в Испании) не может быть определен точно. В другом локусе генотип du du дает окраску в виде белого пояса Голландского кролика.
Мутации структуры волос. Три главных:
- Ангорская. Это - рецессивная аутосомальная мутация, выраженная в удлинении продолжительности роста волос при той же самой скорости роста тела. Дикий (L доминирующий) ген мутировал в рецессивную l аллель, чтобы получить Ангорского кролика. Спаривание двух Ангорских кроликов всегда приводит к получению Ангорского потомства. Два кролика с нормальными волосами могут иногда давать Ангорское потомство, если они Ll гетерозиготы.
- Рекс. Это рецессивная аутосомальная мутация, которая заставляет почти все остевые волосы исчезнуть. Вид меха кролика Рекс отличен от меха нормальношерстных кроликов, поскольку имеет более короткие волосы. Ген короткошерстности обозначается - r, а для доминирующей дикой аллели - R.
- Безволосая. Отсутствие волос. Это вызвано несколькими рецессивными мутациями и обычно смертельно.
Генотип цвета и структуры волосяного покрова пород кроликов может быть предсказан, если локусы исходных животных известны. Пока известно не много генов, явно воздействующих на цвет тела и характеристики размножения, но в этой области было сделано небольшое количество исследований. Ангорский ген и рекс-ген, конечно используются для получения ангорской шерсти и рекс-меха.
Группы пород по размеру взрослых животных и происхождению
Существуют различные виды пород:
- примитивные или первичные породы, и географические, от которых были получены все другие породы;
- породы, полученные путем искусственного отбора первичной породы, такие как Палевый Бургундский, Шампань, Новозеландская Белая и Красная;
- искусственно выведенные породы, полученные плановым скрещиванием нескольких пород, такие как Белый Бускат и Калифорнийский;
- Мендельские породы, полученные фиксацией нового признака простым генетическим закреплением. Появляются в результате мутаций, такие как Кастор-Рекс, Сатиновый и Японский.
Породы удобно сгруппированы по размеру взрослых животных, который также связан с такими производственными характеристиками как раннее развитие, плодовитость, темпы роста и развития в зрелости. Главный детерминант пород взрослого размера - происхождение породы.
Крупные породы. Взрослый вес превышает 5 кг. Продуктивность, главным образом, низкая. Потенциал роста крупных пород может использоваться преимущественно при межпородном скрещивании. Примеры крупных пород: Бускатный Белый Гигант, Французский Баран, Фламандский Гигант и Французская Гигантская Бабочка. Окраска меха Французского Барана сильно варьирует и может быть белой, агути, железистой, серой или черной. Строение тела Французского Барана предусматривает хорошие мясные качества. Однако почти всегда он разводится для показа на выставках и поэтому выращивается только на маленьких фермах, по крайней мере, во Франции. Эта порода более важное значение имеет в других Европейских стран, таких как Германия и Дания.
Бускатный Белый Гигант - искусственная альбиносная порода. Это крупный кролик, известный своей плодовитостью и быстрыми темпами роста в традиционных французских крольчатниках. Фламандский Гигант из Бельгии имеет несколько окрасов. Это один из самых крупных кроликов (потенциальный взрослый вес 7 кг), который все еще выращивается на фермах. Эта порода использовалась в качестве улучшающей по показателю скорости роста при создании новых или улучшению имеющихся пород; Фламандский Гигант мог быть чистокровным для этой цели.
Средние породы. Взрослый вес варьирует от 3.5 до 4.5 кг. Это основные сельскохозяйственные породы, используемые для интенсивного воспроизводства кроликов на мясо в Западной Европе и наиболее многочисленные. Ниже описаны лишь несколько представителей этой группы.
Серебристые кролики обнаружены в нескольких странах (Английский Серебристый, Немецкий Серебристый). Эти разновидности отличаются от породы Шампань размером взрослых особей (Английский Серебристый мельче) и цветом волосяного покрова. Как и Палевый Бургундский, Шампань - пример породы, которую вывели путем отбора и подбора в течение многих лет в жители предместья во Франции (Шампань). Порода известна и за свой мех, и за свою производительность: высокая плодовитость, скороспелость, хорошо развитая мускулатура и хорошее качество мяса. Взрослый вес 4 - 4.5 кг. Она выращивается на фермах во Франции, обычно на соломенной подстилке. Начаты исследования по интенсивному выращиванию породы Шампань.
Палевый Бургундский имеет также региональное происхождение. Уже затем эта порода распространилась по всей Франции и другим странам Европы (Италия, Бельгия, Швейцария). Ассоциация Селекционеров Кролика породы Палевый Бургундский основала племенную книгу для этой породы, гарантируя чистокровность породы.
Новозеландская Красная впервые выведена в Калифорнии методом отбора и подбора. Подобные методы использовали во Франции при выведении породы Палевый Бургундский, но различие в том, что Новозеландская порода была выведена в клетках с сетчатым полом. В Соединенных Штатах сетчатые клетки начали применяться для выращивания кроликов намного раньше, чем во Франции.
Калифорнийская - искусственно выведенная Американская порода. Впервые она была представлена своим создателем в Калифорнии в 1928. Целью этого селекционера было получить животное для производства мяса с очень хорошим мехом. Взрослый вес кроликов породы Калифорнийская - 3.6 - 4 кг.
Новозеландская Белая выведена как порода в Соединенных Штатах. Это потомство альбиносов, полученное от цветных кроликов. Сначала они разводились на нескольких больших фермах по производству мяса, расположенных в южной Калифорнии (в районе г. Сан-Диего). Там они показали хорошие производственные показатели: плодовитость, материнские качества, быстрые темпы роста и скороспелость, которые позволили производить убой молодняка на мясо в возрасте 56 дней и получать молочные тушки. Взрослый вес кроликов породы Новозеландская Белая достигает 4 кг, что слегка превышает тот же показатель у породы Калифорнийская. Новозеландская Белая как порода кроликов впервые была изучена на селекционной станции Фонтана в Калифорнии. С 1960 эта порода распространилась через Западную Европу в другие страны, где для содержания кроликов все больше используют клеточные батареи.
Большая Шиншилла выведена в Европе и имеет немецкое происхождение. Ее взрослый вес составляет в среднем 4.5 кг. Она может разводиться как для мяса, так и для меха.
Мелкие породы. Эти породы имеют взрослый вес 2.5 - 3 кг. К мелким породам относят: Малый Гималайский, Малая Шиншилла, Голландская и Французская Гавана.
Русский или Гималайский кролик имеет белый окрас с черными конечностями. Скорее всего, он был выведен в Китае, а оттуда распространился по России и Польше. Он несет гималайский ген мутации Ch.
Мелкие породы обычно очень быстро развиваются, а самки обладают превосходными материнскими качествами. Они потребляют меньше корма, чем средние и крупные породы и могут использоваться при скрещиваниях или в чистом виде в развивающихся странах для производства некрупных мясных тушек 1 - 1.2 кг.
Карликовые породы. Взрослые животные этих пород весят приблизительно 1 кг. Эта группа представлена в основном Польским кроликом и его многочисленными разновидностями окраса. Селекция, ориентированная на небольшой размер тела кроликов привела к очень низкой плодовитости, и уменьшению скорости роста. Эти породы не могут использоваться для производства мяса. Они разводятся для участия в выставках, для лабораторных целей и в качестве домашних питомцев.
Местные популяции и линии
Чистокровные животные обычно выращиваются в маленьких группах, и их селекция, начиная с рождения, направлена на характеристики породы. Поэтому эти породы могут составлять интересный генофонд (совокупность наследственных факторов популяции) с большим потенциалом для улучшения местных популяций.
Большинство кроликов выращиваемых для промышленного производства мяса, принадлежат к популяциям, которые могут напоминать ту или иную породу (это объясняется тем, что местные кролиководы не встречали критерии оценки той или иной специфической породы в стандартах пород) или иногда не напоминают ни одну из существующих стандартных породу. Это "обычные" кролики, серые, разноцветные или белые, результат различных незапланированных скрещиваний. Они могут принадлежать к местным популяциям. Несколько примеров местных популяций в развивающихся странах - кролик Балади в Судане (baladi, в переводе с арабского – аборигенный, или местный), Мальтийский кролик в Тунисе, Креольский кролик в Гваделупе. Развивающиеся страны, планирующие развивать промышленное производство кролика должны сначала идентифицировать существующие местные популяции и установить их биологические особенности и особенности размножения, а также адаптируемость до вступления в силу селекционных программ и улучшенных систем производства.
Многие страны, где выращиванием кроликов занимаются не так давно, порядка несколько десятилетий, имеют не четко определенные местные популяции. Популяции высоко полиморфичны и получены от очень многих незапланированных скрещиваний с импортированными чистокровными животными. Часто эти популяции имеют ограниченный потенциал и не приспособлены к местным условиям. Даже в этом случае, они должны быть изучены, даже если принято решение устранить их.
Наконец, существуют линии кроликов. Линия - не многочисленная генетически закрытая группа, воспроизводимая внутри себя, т.е. без применения скрещиваний извне, на протяжении нескольких поколений. Характеристики линии: количество животных участвовавших в создании линии, количество лет выведения и метод по которому группа была составлена, и возможно программа спариваний (селекция или ее отсутствие). Эти линии могут быть найдены в научно-исследовательских лабораториях, которые продолжают изучать их биологические характеристики и характеристики размножения, чтобы сделать их лучше для использования в целях селекции. Центр INRA в Тулузе сосредоточен на селекционных экспериментах с линиями (табл. 28).
Частные селекционеры довольно недавно начали селекционировать линии кроликов, подобно линиям селекции домашней птицы, селекция которых были осуществлена еще с 1930. Но некоторые селекционеры или небольшие группы селекционеров, на деревенском уровне сами того не осознавая, создали линии.
Некоторые научно-исследовательские лаборатории, типа Лаборатории Джексона в Бар Гарборе, штат Мэн, Соединенные Штаты, содержат инбредные линии кроликов или линии для использования исключительно в качестве лабораторных животных.
Селекционеры в районах традиционно выращивающих кроликов используют местные популяции. Генетическое наследие популяции сформировано местной экологией, характерной системой производства и вмешательствами кролиководов. Постепенно, популяция развивается. За исключением определенных случаев, популяция открыта для приграничных популяций. Это замедляет тенденцию достижения однородности и оставляет возможность введения новой генетической изменчивости для естественной и/или искусственной селекции.
Следующая стадия развития – порода. Здесь селекционер более важен, поскольку он определяет стандарт породы, и методом отбора и подбора животных стремится к соответствию стандарта. Экология региона и характеристики производственной системы в меньшей степени влияют на популяцию, а породы обычно более однородны по этим критериям. Стремление селекции приблизиться к идеальному стандарту породы может привести к выходу за пределы допустимого. Для достижения цели селекционеры могут оценивать только внешние признаки, пренебрегая показателями продуктивности. Они могут применять скрещивания между близкими родственниками, чтобы увеличить визуальное впечатление от однородности. Последний эволюционный уровень - линия. Она имеет меньшее количество потомков (несколько дюжин каждого пола) и небольшое количество генов заимствованных у смежных популяций. Линия обычно искусственно селекционируется по нескольким параметрам. Часто линии генетически более однородны, чем породы.
Характеристики размножения
Выражение характеристик размножения зависит от окружающей среды и селекционера. Сравнение результатов экспериментов, проводимых в различных окружающих средах и географических зонах, может показать общие характеристики пород или видов. Плодовитость, скорость роста и развития тканей у молодняка кроликов - три основные характеристики размножения.
Плодовитость. Общая плодовитость определяется как результат фертильности (количество окролов на самку за единицу времени) и плодовитости за окрол (количество крольчат в окроле).
Плодовитость значительно изменяется под воздействием некоторых факторов, которые могут быть свойственны животному. Размер помета второго окрола увеличивается на 10 - 20% по сравнению с первым, а затем от второго к третьему снова уменьшается, от третьего до четвертого окрола количество крольчат в окроле остается без изменения. После четвертого окрола размер помета может уменьшаться. Инбридинг также может уменьшать плодовитость.
ТАБЛИЦА 28 Характеристики экспериментальных линий селекционированных INRA
 Плодовитость также зависит от времени года и репродуктивной нормы, возложенной на самку. У здоровых самок, получающих нормальный корм, освещение по 12 - 14 часов в сутки, сниженная плодовитость может быть связана с ненормальным весом самок.
Плодовитость также зависит от времени года и репродуктивной нормы, возложенной на самку. У здоровых самок, получающих нормальный корм, освещение по 12 - 14 часов в сутки, сниженная плодовитость может быть связана с ненормальным весом самок.
Количество овулировавших яйцеклеток увеличивается соответственно с размером тела самки. Основной фактор, влияющий на плодовитость - степень овуляции (количество яйцеклеток овулировавших одновременно), далее, вплоть до рождения, на плодовитость влияет уровень жизнеспособности бластоцист и эмбрионов.
В 1932, Gregory доказал, что размер помета зависит от количества овулировавших после спаривания яйцеклеток, и это количество зависит от размера тела каждой породы: 3.97 для крольчихи породы Польский и 12.88 для Фламандского Гиганта. Размеры помета при рождении составляют 3.24 и 10.17 соответственно. Вообще, мелкие легкие породы менее плодовиты, чем средние и большие породы. Elamin (1978) дает следующие средние показатели по Судану для кроликов дующих пород: Балади, Калифорнийская и Новозеландская Белая:
 Matheron и Dolet (1986) проанализировали результаты, полученные от 682 самок из десяти крольчатников на Гваделупе. Первая категория - Креольские самки небольшого размера. Их трудно найти, так что селекционеры покупали их во Франции, а затем в течение длительного времени скрещивали их с различными породами кроликов. Затем они разделили самок на Новозеландских Белых и "других" самок, т.е. всех тех, по которым данные не достаточно определенные. В этих комплексных скрещиваниях, селекционеры использовали породы: Шампань, Палевый Бургундский, Бускатный Белый Гигант, Бабочка и ещё две линии. Табл. 29 показывает, что Новозеландские самки более фертильные и более плодовитые. Это подтверждает высокую адаптацию этой породы к местным условиям содержания, отличающимся высокой температурой и влажностью. Смертность в периоде от рождения до отъема все еще высока, что указывает на потребность дальнейшего изучения этого вопроса. Креольские самки менее плодовиты, но более жизнеспособны чем "другие" самки. При рождении расхождение - 0.78, а при отъеме только - 0.12. Слабая жизнеспособность молодняка в промежутке от рождения до отъема у "других" самок удивляет. Литература указывает, что эти помесные самки часто извлекают выгоду из эффекта гетерозиса и взаимодополняемости, но представленная работа показывает, что это не всегда является правилом, а лишь дело случая. Также возможно, что выбор пород и кроссов был недостаточен.
Matheron и Dolet (1986) проанализировали результаты, полученные от 682 самок из десяти крольчатников на Гваделупе. Первая категория - Креольские самки небольшого размера. Их трудно найти, так что селекционеры покупали их во Франции, а затем в течение длительного времени скрещивали их с различными породами кроликов. Затем они разделили самок на Новозеландских Белых и "других" самок, т.е. всех тех, по которым данные не достаточно определенные. В этих комплексных скрещиваниях, селекционеры использовали породы: Шампань, Палевый Бургундский, Бускатный Белый Гигант, Бабочка и ещё две линии. Табл. 29 показывает, что Новозеландские самки более фертильные и более плодовитые. Это подтверждает высокую адаптацию этой породы к местным условиям содержания, отличающимся высокой температурой и влажностью. Смертность в периоде от рождения до отъема все еще высока, что указывает на потребность дальнейшего изучения этого вопроса. Креольские самки менее плодовиты, но более жизнеспособны чем "другие" самки. При рождении расхождение - 0.78, а при отъеме только - 0.12. Слабая жизнеспособность молодняка в промежутке от рождения до отъема у "других" самок удивляет. Литература указывает, что эти помесные самки часто извлекают выгоду из эффекта гетерозиса и взаимодополняемости, но представленная работа показывает, что это не всегда является правилом, а лишь дело случая. Также возможно, что выбор пород и кроссов был недостаточен.
Paez Campos и др. (1980) дают параметры размножения таких пород как Новозеландская Белая, Калифорнийская, Шиншилла, и Рекс разводимых в Национальном Центре Кролиководства Ирапуато в Мексике, расположенного в тропической зоне на высоте 1 800 м над уровнем моря (табл. 30).
ТАБЛИЦА 29 Показатели продуктивности самок трех генетических типов в крольчатниках Гваделупы
 Источник: Matheron и Dolet, 1986.
Источник: Matheron и Dolet, 1986.
ТАБЛИЦА 30 Средние показатели продуктивности четырех пород, выращиваемых в Национальном Центре Кролиководства Ирапуато, Мексика
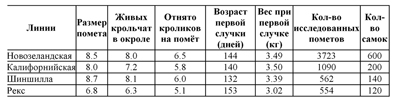 ТАБЛИЦА 31 Показатель размера помета по четырем породам кроликов на Кубе
ТАБЛИЦА 31 Показатель размера помета по четырем породам кроликов на Кубе
 Источник: Ponce de Léon, 1977
Источник: Ponce de Léon, 1977
ТАБЛИЦА 32 Резюме селекционируемых пород сравниваемых по индивидуальному весу при отъеме, индивидуальному весу в 10 недель, размеру гнезда при окроле и отъеме
 Ponce de Léon (1977) получил результаты, представленные в табл. 31 по четырем породам, исследуемым на Кубе, в условиях влажного тропического климата. Характеристики этих пород и этих крольчатников более детально описаны ниже. Высокий уровень смертности (11.6%) объясняется условиями содержания в крольчатнике.
Ponce de Léon (1977) получил результаты, представленные в табл. 31 по четырем породам, исследуемым на Кубе, в условиях влажного тропического климата. Характеристики этих пород и этих крольчатников более детально описаны ниже. Высокий уровень смертности (11.6%) объясняется условиями содержания в крольчатнике.
Развитие технических и экономических производственных систем во Франции и Испании обеспечивает ряд результатов, описывающих тенденции изменения производительности в промышленных крольчатниках. Для примера, систематическая работа в Институте технического оснащения кролико… Продолжение »